Ceruloplasmin: The Master Regulator of Iron
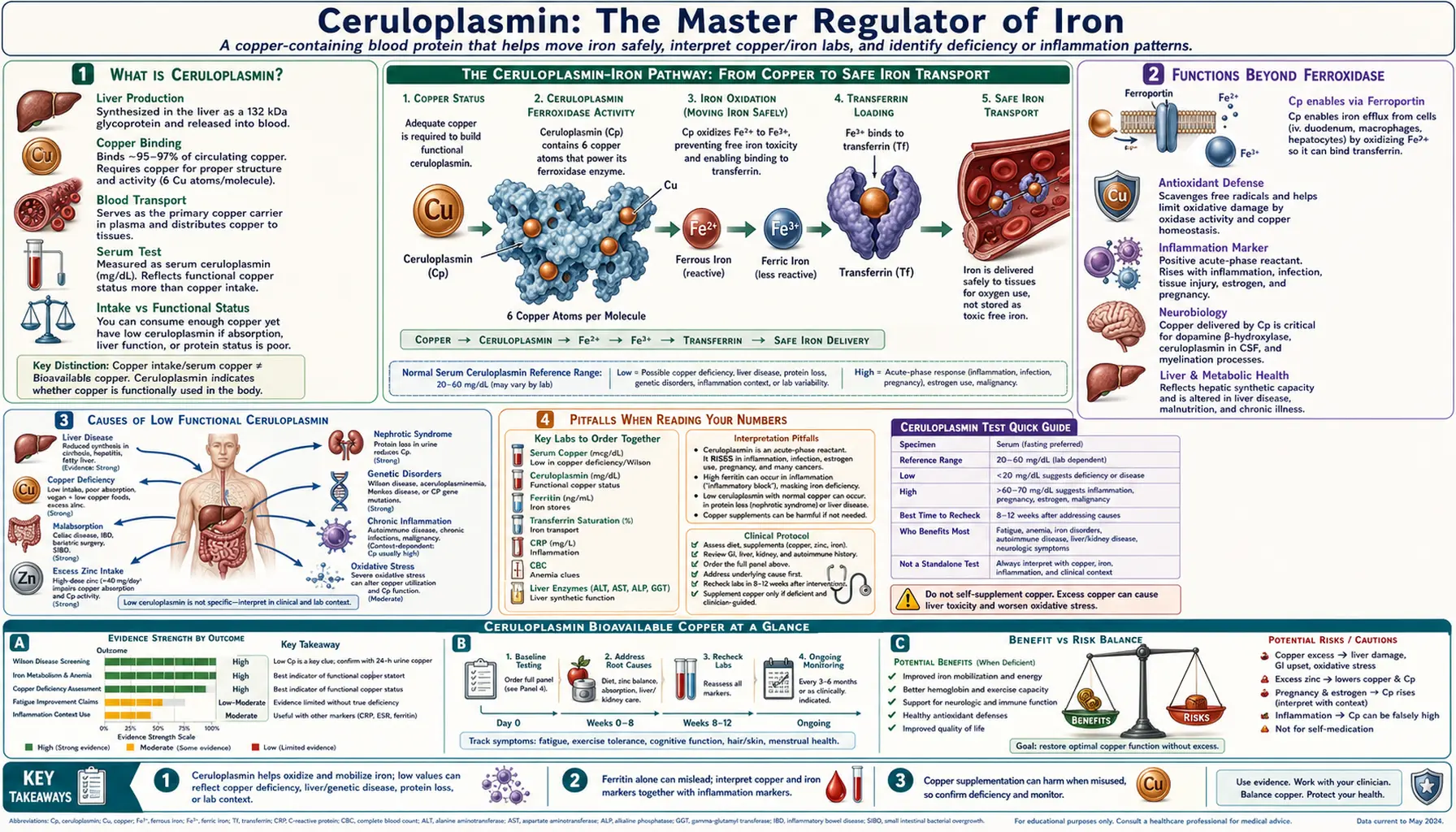
Of all the proteins in human plasma, none has been more central to Morley Robbins’s teaching — or more underappreciated by mainstream lab interpretation — than ceruloplasmin. It is the protein the Root Cause Protocol returns to again and again, the master regulator of iron metabolism, the carrier of nearly all circulating copper, and the single number Robbins says he would look at first if he could only pick one lab. This article is a deep dive into what ceruloplasmin is, what it does, how to read it on a standard panel, the “bioavailable copper” calculation Robbins teaches his clients, the causes of low functional ceruloplasmin, and how to raise it through whole-food and lifestyle change. Where mainstream physiology agrees with the RCP framing, we say so; where it diverges, we say that too.
Table of Contents
- What Is Ceruloplasmin?
- The Six Copper Atoms: Structure and Activity
- Functions Beyond Ferroxidase
- Lab Reference Ranges and What “Normal” Hides
- The Bioavailable Copper Calculation
- Reading the Full Lab Panel
- Causes of Low Functional Ceruloplasmin
- How to Raise Ceruloplasmin
- Pitfalls When Reading Your Numbers
- Key Research Papers
- Connections
- Featured Videos
1. What Is Ceruloplasmin?
Ceruloplasmin is a 132-kilodalton multi-copper oxidase synthesized in the liver by hepatocytes and secreted into plasma. It is the principal copper-carrying protein in blood, transporting roughly 95% of all circulating copper. The remaining 5% is loosely bound to albumin, the amino acid histidine, and a small amount of low-molecular-weight peptides — this fraction is what Robbins and the laboratory medicine literature call the “free,” “non-ceruloplasmin,” or “bioavailable” copper.
The plasma half-life of ceruloplasmin is approximately 5.5 days, which means a measurable shift in the lab number can take a week or more to register after a dietary or supplement change. The protein is glycosylated, which gives it a pale blue color when concentrated — the name “ceruloplasmin” literally means “sky-blue plasma protein,” from the Latin caeruleus.
Ceruloplasmin is synthesized inside the hepatocyte first as apoceruloplasmin — the protein backbone without any copper attached. Loading the six copper atoms into the protein is performed by the ATP7B copper-transporting ATPase, a transmembrane pump in the trans-Golgi network of the hepatocyte. ATP7B is the same gene that, when mutated, causes Wilson’s disease, a hereditary copper-overload disorder. In Wilson’s disease, copper cannot be loaded into apoceruloplasmin or excreted into bile, so the hepatocyte accumulates copper to toxic levels while plasma ceruloplasmin runs low — the classic Wilson’s pattern of low ceruloplasmin with high free copper.
The flip side is also clinically important: apoceruloplasmin without its copper cargo is rapidly degraded. So a copper-poor liver makes less functional ceruloplasmin even at the same level of gene expression. This is why “take more zinc to lower copper” protocols can paradoxically crash ceruloplasmin — the gene is firing fine, but there is no copper to load, and the empty apoprotein is destroyed.
Ceruloplasmin was first isolated and named in 1948 by Carl-Bertil Laurell and Curt-G. Holmberg in Sweden. Its ferroxidase activity — the ability to oxidize ferrous iron (Fe²⁺) to ferric iron (Fe³⁺) so that it can be loaded onto transferrin — was characterized by Earl Frieden and colleagues in the 1960s. The 2002 review by Hellman and Gitlin in Annual Review of Nutrition remains the definitive modern synthesis of ceruloplasmin biology.
2. The Six Copper Atoms: Structure and Activity
Each ceruloplasmin molecule contains exactly six copper atoms bound at three structurally and functionally distinct sites. Understanding the geometry helps explain why the protein is both an iron-oxidizing enzyme and a copper carrier:
- Type I “blue” copper sites — three coppers. These give the protein its characteristic color. They are coordinated by cysteine and histidine residues and serve as the primary electron-acceptor sites. Substrate molecules bind near these centers and donate an electron to copper, beginning the catalytic cycle.
- Type II copper site — one copper. Coordinated by histidines, this center sits adjacent to the Type III pair and participates in the trinuclear oxygen-reducing cluster.
- Type III copper pair — two coppers. A pair of coppers held in close proximity (~3.5 Å apart) and bridged by a hydroxide ion. Together with the Type II copper, this trinuclear cluster performs the four-electron reduction of molecular oxygen (O₂) to water (H₂O) during the oxidation of ferrous iron.
The trinuclear cluster is what gives ceruloplasmin its ferroxidase activity. Four ferrous iron substrates each donate one electron to the Type I sites, the electrons are passed through the protein to the trinuclear cluster, and there they reduce a single O₂ molecule to two H₂O molecules. Net: 4 Fe²⁺ + O₂ + 4 H⁺ → 4 Fe³⁺ + 2 H₂O. The cleanly reduced water output is critical — it means iron oxidation by ceruloplasmin produces no hydroxyl radicals, in stark contrast to the uncatalyzed Fenton-reaction iron oxidation that drives oxidative tissue damage.
This is the molecular core of Robbins’s argument: ceruloplasmin is the body’s safe-iron-oxidation machine. Without enough functional ceruloplasmin, ferrous iron in tissues is left to undergo Fenton chemistry with hydrogen peroxide, generating the hydroxyl radicals that damage lipids, proteins, and DNA.
3. Functions Beyond Ferroxidase
Although ferroxidase is the headline activity, ceruloplasmin is a multifunctional protein:
- Ferroxidase activity. Oxidation of Fe²⁺ to Fe³⁺ for loading onto transferrin. This is essential for delivering iron from intestinal absorption, macrophage iron recycling, and hepatic iron stores into the plasma transport pool. The closely related membrane protein hephaestin performs the analogous function at the basolateral surface of the enterocyte.
- Amine oxidase activity. Ceruloplasmin oxidizes biogenic amines including serotonin, norepinephrine, dopamine, and various dietary amines. This is one reason copper status influences mood, sleep, and stress physiology — ceruloplasmin participates in clearing the same neurotransmitters that govern those systems.
- Glutathione peroxidase-like activity. A secondary antioxidant role: ceruloplasmin can quench certain reactive oxygen species and prevent lipid peroxidation in plasma.
- Copper transport. Although ceruloplasmin is no longer thought to be the primary copper-delivery vehicle to peripheral tissues — that role is now attributed to albumin and amino-acid-bound copper, particularly histidine-bound copper — ceruloplasmin does deliver copper to certain cell types and is the major plasma copper reservoir.
- Indirect role in cholesterol homeostasis. Through its regulation of iron metabolism and its amine-oxidase activity, ceruloplasmin participates indirectly in lipid biology. Low ceruloplasmin states have been associated with altered cholesterol handling in several observational studies.
- Innate immune function. Ceruloplasmin is an acute-phase reactant — it rises during infection — and contributes to the body’s strategy of withholding iron from invading microbes (“nutritional immunity”).
4. Lab Reference Ranges and What “Normal” Hides
The standard laboratory reference range for serum ceruloplasmin is roughly 20–35 mg/dL in adults, with some labs quoting 18–45 mg/dL. Reference ranges vary slightly by assay method and by lab.
Robbins’s position is that anything below 30 mg/dL represents functional copper deficiency, and his RCP target for clients is around 35 mg/dL. He argues that the low end of the standard reference range was set by sampling a population already deficient — that is, “normal” reflects the unhealthy modern average, not an optimal physiological set-point. Mainstream laboratory medicine sets the low cutoff for clinical concern at 20 mg/dL, which is where Wilson’s disease workup is typically triggered.
Several physiological states alter ceruloplasmin without any pathology:
- Pregnancy. Ceruloplasmin roughly doubles by the third trimester, with values of 40–60 mg/dL being normal in pregnancy. This is driven by estrogen, which is a strong inducer of hepatic ceruloplasmin gene transcription.
- Estrogen-containing oral contraceptives raise total ceruloplasmin similarly — sometimes by 50–100% — without necessarily raising functional ferroxidase activity proportionally.
- Acute phase response. Ceruloplasmin rises in infection, inflammation, malignancy, and trauma. A value in the “adequate” range during an acute illness may mask an underlying deficiency.
- Newborns. Newborns have very low ceruloplasmin (5–10 mg/dL) that rises rapidly over the first 6–12 months of life.
Causes of low ceruloplasmin include copper deficiency, severe liver disease, Wilson’s disease (low CP with high free copper, plus elevated 24-hour urinary copper), Menkes disease (genetic CP synthesis defect), aceruloplasminemia (rare absence of CP), nephrotic syndrome (urinary protein loss), and severe malabsorption syndromes.
5. The Bioavailable Copper Calculation
The single most useful quantitative tool in Robbins’s lab-reading toolkit is the bioavailable copper calculation. The math is simple, but the conceptual move is powerful: it forces you to look at the relationship between total copper and ceruloplasmin-bound copper, rather than at either number in isolation.
Step 1. Convert ceruloplasmin (mg/dL) into the copper it actually carries (µg/dL) by multiplying by 3:
Ceruloplasmin-bound copper (µg/dL) = Ceruloplasmin (mg/dL) × 3
The conversion factor of 3 derives from the protein’s structure. Ceruloplasmin carries 6 copper atoms per molecule. Copper has an atomic mass of 63.5 g/mol. Ceruloplasmin’s molecular weight is 132,000 g/mol. The fraction of ceruloplasmin mass that is copper is therefore (6 × 63.5) / 132,000 ≈ 0.00289, or roughly 0.3%. So each milligram of ceruloplasmin carries about 3 micrograms of copper.
Step 2. Subtract ceruloplasmin-bound copper from total serum copper to get the bioavailable (“free” or “non-ceruloplasmin”) copper:
Bioavailable copper (µg/dL) = Serum copper (µg/dL) − Ceruloplasmin-bound copper (µg/dL)
Step 3. Express bioavailable copper as a percentage of total copper:
% Bioavailable = (Bioavailable copper / Serum copper) × 100
Healthy reference for the ratio. The mainstream Wilson’s-disease threshold is greater than 25% free copper, which is considered diagnostic when accompanied by other findings. Robbins’s functional target is roughly 10–25% bioavailable — meaning most of the circulating copper should be safely loaded into ceruloplasmin, with a small reserve in the “free” fraction for delivery to tissues.
Worked example. A client presents with serum copper = 120 µg/dL and ceruloplasmin = 20 mg/dL.
- Ceruloplasmin-bound copper = 20 × 3 = 60 µg/dL
- Bioavailable copper = 120 − 60 = 60 µg/dL
- % bioavailable = 60 / 120 × 100 = 50%
Both the total serum copper (120) and the ceruloplasmin (20) might individually be flagged as “within range” by some lab cutoffs. But 50% bioavailable copper is well above the 25% Wilson’s threshold and reflects a state in which half of the body’s circulating copper is not safely held in its ferroxidase carrier protein. Robbins distinguishes between enzymatically-active copper (bound to ceruloplasmin and other functional cuproenzymes) and unbound copper, which may be reactive and contribute to oxidative stress. The latter is what the high-percent-bioavailable result is highlighting.
6. Reading the Full Lab Panel
Ceruloplasmin and serum copper alone are not enough. Robbins teaches a standard order list (the “Full Monty” in his terminology — an unrelated American expression he uses for the complete diagnostic picture) so the copper–iron axis can be evaluated as a whole:
- Serum copper (µg/dL)
- Ceruloplasmin (mg/dL)
- Ferritin (ng/mL) — the inflammation/iron-storage marker; Robbins targets 20–80 in women and 30–90 in men, far below the lab high-cutoff of 200–400
- Iron, total serum (µg/dL)
- TIBC — total iron binding capacity (µg/dL)
- Transferrin saturation % (target 25–45%)
- Hemoglobin (g/dL), hematocrit (%), and red-cell indices (MCV, MCH, MCHC)
- A/G ratio (albumin/globulin), drawn from a comprehensive metabolic panel
- RBC magnesium (preferred to serum magnesium, which is poorly representative of tissue stores)
- 25-OH vitamin D and active 1,25-(OH)₂ vitamin D (pair them — an isolated 25-OH-D is incomplete)
- Retinol (serum, ideally separated from beta-carotene)
- Zinc (RBC zinc preferred to serum zinc)
The relationships matter as much as the absolute numbers. Low ceruloplasmin with normal serum copper means high free copper. High ferritin with low transferrin saturation and low iron suggests inflammation rather than true iron stores. Low retinol with low ceruloplasmin suggests the cofactor for CP synthesis is missing. The patterns drive the protocol — not single out-of-range flags.
7. Causes of Low Functional Ceruloplasmin
Robbins’s teaching catalogues a long list of factors that suppress ceruloplasmin synthesis or accelerate its degradation. Several are uncontroversial in mainstream physiology; others are RCP-specific claims supported by mechanistic reasoning more than by RCT evidence.
- Dietary copper deficiency. Modern processed diets, soil depletion, and vegetable-heavy diets without organ meat or shellfish are common contributors.
- Retinol (vitamin A) deficiency. Retinoic acid is required for transcription of the ceruloplasmin gene. Beta-carotene from plants is converted to retinol with poor and variable efficiency, so plant-only sources may be inadequate.
- High-dose zinc supplementation. Zinc induces metallothionein in enterocytes, which sequesters copper and prevents absorption. Long-term zinc supplementation above ~50 mg/day is a recognized cause of functional copper deficiency.
- High-dose synthetic ascorbic acid. Some studies suggest gram-dose ascorbic acid lowers ceruloplasmin; the literature is mixed and the mechanism debated. Robbins favors whole-food vitamin C from acerola or camu camu over synthetic ascorbic acid.
- High-dose iron supplementation. Iron and copper compete for absorption in the enterocyte; chronic iron supplementation can drive functional copper deficiency.
- High-dose vitamin D3 supplementation. Per Robbins, cholecalciferol depletes magnesium and retinol — both required for ceruloplasmin biology — and suppresses ceruloplasmin. The mechanism is debated and the magnitude is dose-dependent (the effect is most plausible at the >10,000 IU/day range, not at typical 1,000–2,000 IU doses).
- Severe liver disease. Cirrhosis and advanced hepatitis impair hepatocyte synthetic capacity.
- Wilson’s disease. Genetic ATP7B mutation prevents copper loading into apoceruloplasmin; the empty apoprotein is degraded, plasma CP runs low, and copper accumulates in the hepatocyte and brain.
- Menkes disease. Genetic ATP7A mutation; failure of intestinal copper absorption and ceruloplasmin synthesis.
- Aceruloplasminemia. Rare genetic absence of ceruloplasmin altogether; produces brain and pancreatic iron overload.
- Estrogen birth control. Paradoxically raises total ceruloplasmin while often lowering bioavailable copper through a “ceruloplasmin trap” effect — copper is bound up in CP but functional ferroxidase activity may not rise proportionally.
- Chronic stress and elevated cortisol. Cortisol’s catabolic effects deplete the cofactor pool over time.
- Glyphosate exposure. RCP teaching points to glyphosate as a chelator of trace minerals including copper and as a disruptor of liver function.
- Fluoride exposure. Per RCP teaching, fluoride disrupts mineral metabolism broadly and lowers ceruloplasmin.
8. How to Raise Ceruloplasmin
The RCP approach to raising ceruloplasmin is whole-food and time-paced. Robbins emphasizes that ceruloplasmin’s 5.5-day half-life and the slower turnover of the cofactor pool mean changes show up over three to six months, not three to six days.
- Whole-food copper sources. Beef liver 1–3 ounces weekly (ideally pastured), oysters, dark chocolate and unsweetened cacao, bee pollen, sesame seeds, cashews, shiitake and crimini mushrooms. The Whole-Food Copper Sources page has the full ranked list.
- Whole-food retinol sources. Cod liver oil (~1 teaspoon per day, fermented or low-heat-processed), pastured egg yolks, butter from grass-fed cows, beef liver. Retinol — not beta-carotene — is what activates ceruloplasmin gene transcription.
- Magnesium. Glycinate, malate, taurate, or threonate forms at 400–800 mg/day. Magnesium is required for the enzymes that activate vitamins A and D and that maintain mitochondrial energy production at the hepatocyte.
- Whole-food vitamin C. Acerola cherry, camu camu, rose hip powder — the goal is the bioflavonoid-and-copper-cofactor matrix that comes with food, not isolated ascorbic acid.
- Stop the “Stops.” Discontinue or minimize industrial iron-fortified flour and cereal, synthetic ascorbic acid, calcium supplements, and high-dose vitamin D3. (See the Root Cause Protocol page.)
- Adrenal cocktail. Robbins’s simple recipe of orange juice, sea salt, and cream of tartar to support adrenal cortisol production and the mineral pool the adrenals draw from.
- Reduce chronic stress. Cortisol burns through the same cofactor pool that supports ceruloplasmin synthesis.
- Sunlight exposure. Sunlight on bare skin produces endogenous vitamin D in concert with the cholesterol pathway, without depleting the magnesium and retinol cofactors that supplemental D3 may consume.
- Time and patience. Three to six months is the realistic timeframe for measurable ceruloplasmin improvement on labs — not weeks.
9. Pitfalls When Reading Your Numbers
Several pitfalls recur often enough that they deserve their own list:
- Acute-phase elevation. Ceruloplasmin rises during infection, inflammation, surgery, trauma, and certain malignancies. A “normal” CP drawn during a sinus infection or after a recent injury may mask underlying deficiency. Wait at least 4–6 weeks after acute illness to draw a baseline.
- Pregnancy and estrogen. Pregnancy and estrogen-containing contraceptives elevate total ceruloplasmin without proportionally raising functional capacity. Bioavailable copper may still be high in those states.
- Immunoreactive vs. enzymatically active assays. Most clinical labs measure ceruloplasmin by immunoassay, which detects both apoceruloplasmin and holoceruloplasmin (the loaded form). Specialized labs can measure ferroxidase activity directly, and the two numbers can diverge significantly — a person with adequate immunoreactive CP but low ferroxidase activity is functionally copper-deficient even though the standard lab looks fine.
- Apoceruloplasmin is biologically inactive. The empty apoprotein still shows up on standard immunoassays. This is part of why the “immunoreactive”-versus-“active” distinction matters.
- Serum copper alone is misleading. A normal total copper with low ceruloplasmin means high free copper — often a worse picture than a uniformly low total. Always pair the two numbers.
- Ferritin alone is misleading. Ferritin rises in inflammation, masking iron deficiency in the inflamed patient and falsely reassuring the iron-overloaded patient. Pair ferritin with transferrin saturation and CRP.
- Don’t over-react to a single draw. Ceruloplasmin can swing 10–20% week to week in healthy people. Trends over multiple draws matter more than a single number.
- Lab method differences. Reference ranges differ slightly between immunoassay platforms; comparing across labs without correction can produce phantom changes.
10. Key Research Papers
- Hellman & Gitlin (2002) — definitive ceruloplasmin review (Annu Rev Nutr)
- PubMed: ceruloplasmin ferroxidase activity
- PubMed: apoceruloplasmin
- PubMed: ATP7B Wilson’s disease
- PubMed: Menkes disease ATP7A
- PubMed: ceruloplasmin pregnancy estrogen
- PubMed: ceruloplasmin acute-phase reactant
- PubMed: copper ceruloplasmin inflammation
- PubMed: hephaestin enterocyte
- PubMed: ceruloplasmin amine oxidase
Connections
- Morley Robbins
- Root Cause Protocol
- Whole Food Copper Sources
- Copper-Iron Dysregulation
- Iron Overload and Hidden Toxicity
- Magnesium Replenishment
- Vitamin D Controversy
- Adrenal Cortisol and Mineral Connection
- Cure Your Fatigue
- Copper
- Iron
- Magnesium
- Zinc
- Vitamin A
- Vitamin C
- Vitamin D3
- Vitamin D Test
- Liver Disease